
Charles R. Warren1
1School of Biological Sciences, The University of Sydney, NSW 2006, Australia.
First Published: ASMS 2010
Introduction
- Metabolomics is providing major insights into what it is that makes plants tolerant of abiotic stresses (e.g. drought).
- To date, plant metabolomics has focused on model species (e.g. Arabidopsis)
- Unfortunately the special traits that make conventional model species easy to work with also makes them non-representative of many plants
- Model species are among the least stress-tolerant plants in the world. In fact, the strategy of model species for coping with stress is to die and/or reproduce
- We need additional models species that are more representative of perennial plants (e.g. trees) and stress-tolerant plants
- Eucalyptus species are obvious choices as models for evergreen trees due to their economic and ecological importance and because the genome of one species is being sequenced
- One problem is that available GC-MS libraries are for model species and do not contain many metabolites that are abundant in stress tolerant species.
Aims of this study were to:
- Develop a GC-MS library of metabolites found in Eucalyptus species
- Use untargeted GC-MS of organic metabolites and targeted CE-UV analyses of inorganic ions to explore how Eucalyptus species cope with prolonged water stress.
Plant material and treatments
- Seedlings of 7 Eucalyptus species were exposed to water stress for 3 months. This poster presents data only for 2 contrasting species
- 5 or 6 plants of each species were assigned to each of a water stress treatment and well-watered control
- Water stress was imposed by letting pots dry slowly over a period of ten days until stomatal conductance was < 0.05 mmol m-2 s-1 ( vs >0.3 in controls).
- This level of stress was maintained for 3 months by watering pots each day to a constant mass
- Leaf discs were collected at the end of the 3-month water stress treatments and a week after re-watering.
Analytical methods
- Aqueous metabolites were extracted with methanol:chloroform:water
- Untargeted analysis was based on GC-(EI)-MS of TMS derivatives.
- EI mass spectra were deconvoluted using AnalyzerPro® and metabolites identified by comparing retention indices and mass spectra with an in-house library plus the Golm Metabolome Database, Agilent Fiehn and NIST libraries.
- The in-house library of 130 chemical standards was analysed under the same GC-(EI)-MS conditions. It included key metabolites not in existing libraries.
- Methane CI was used to confirm identifications and provide information aboutunknowns (e.g. MH+)
- Amino acids were quantified by GC-(EI)-MS as t-BDMS derivatives. Inorganic anions and inorganic cations were quantified by capillary electrophoresis with indirect UV detection
Library development
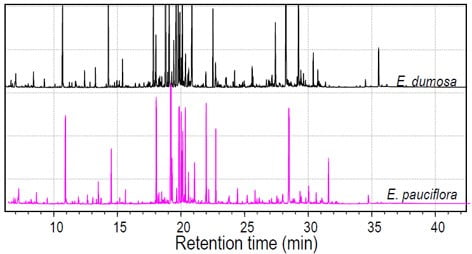
Figure 1. Typical GC-(EI)-MS chromatograms of TMS derivatives of the aqueous fraction of a methanol:chloroform:water extracts from leaves of two species of Eucalyptus
In-house GC-(EI)-MS library for Eucalyptus species
- 180 compounds identified (vs. standards, NIST, Fiehn and Golm libraries)
- 10 match unknowns in Golm database
- 15 unknowns not in any database
In-house GC-(CI)-MS library:
- > 100 metabolites
- Used for confirming EI identifications
- Picking up compounds missed by deconvolution of EI spectra, e.g. probing chromatograms with characteristic ions for different chemical classes (Fig 4)
Untargeted approach uncovers dichotomy in abundant metabolites
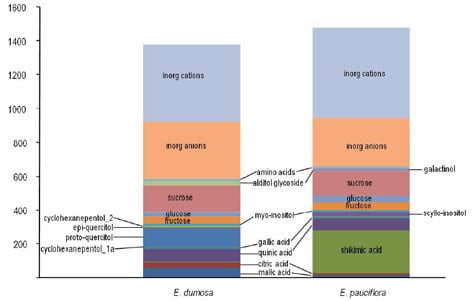
Figure 2. Concentration (in mmol kg leaf water-1) of major solutes in leaves of Eucalyptus dumosa and Eucalyptus pauciflora. Proto-quercitol in E. dumosa is 1.1% of dry mass and shikimic acid in E. pauciflora is 2.9% of dry mass. Only 2 species are shown here, but this dichotomy between occurrence of cyclohexanpentols versus shikimic acid is consistent among 7 Eucalyptus spp.
- Previous targeted analyses with Eucalyptus have shown a dichotomy between those species that contain large concentrations of the cyclohexanepentol proto-quercitol and those that do not (e.g. Merchant et al. Phytochemistry 67: 402-408)
- Untargeted approach adds an extra dimension by showing absence of cyclohexanepentols is associated with extremely high amounts of shikimic acid (3-5% of dry mass)
- Probing methane-CI chromatograms of cyclohexanepentol-containing species (Fig 3 and 4) showed that in addition to large amounts of protoquercitol, cyclohexanepentol-containing species contain smaller amounts of vibo-quercitol, epi-quercitol and three unidentified cyclohexanepentol isomers.
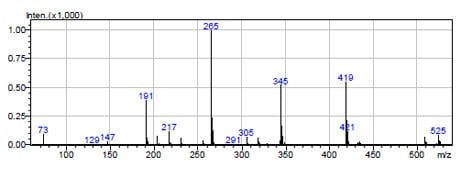
Figure 3. methane CI mass spectrum of proto-quercitol. MH+ = 525
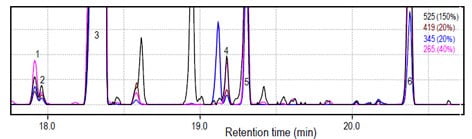
Figure 4. GC-(CI)-MS chromatogram of E. dumosa with ions characteristic ofcyclohexanepentols. 1,2,6 unknown; 3 = proto-quercitol, 4 = epi-quercitol; 5=viboquercitol
Complex and species-specific effect of drought on metabolites
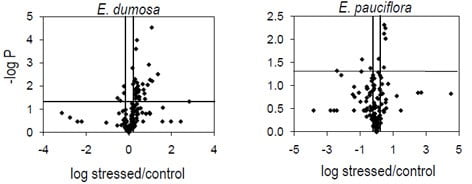
Figure 5. Volcano plots showing effect of severe water stress on leaf metabolites. Metabolites above the horizontal line are statistically significant (P = 0.05), while those to the left and right of the vertical lines are biologically significant (= 1.5-fold difference)
- In E. dumosa 31 metabolites were present at larger concentrations in waterstressed plants than controls, while four were present at smaller concentrations
- In E. pauciflora only 6 metabolites were present at larger concentrations in water-stressed plants than controls, and 3 at smaller concentrations in waterstressed plants than controls
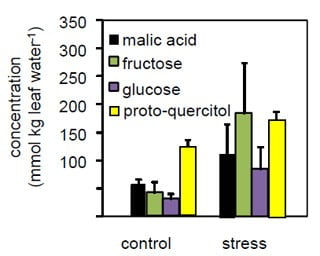
Figure 6. Column graph showing those metabolites in E. dumosa with the largest absolute increases due to water stress.
Conclusions
Confirmed results of previous studies:
- dichotomy between Eucalyptus spp. with and without cyclohexanepentols
- aqueous fraction dominated by cyclohexanepentols, mono- and di-saccharides, and organic acids
- water stress increases Frc, Glc (and in some species Suc)
Added extra dimensions:
- species without cyclohexanepentols have huge concentrations of shikimic acid
- Eucalyptus spp. contain at least 6 (of 16) possible cyclohexanepentol stereoisomers
- water stress affects many monosaccharides, sugar alcohols and cyclohexanepentols
- responsiveness of aqueous metabolites to water stress differs between spp.
Acknowledgements
This work was supported by a Discovery Grant and QEII Fellowship from the Australian Research Council, and a major equipment grant from The University of Sydney. Dr Maria Taranto is thanked for assisting with preparation of samples.