
A GC-MS-based longitudinal untargeted metabolomic analysis of plasma from the Lewis Polycystic Kidney rat model of nephronophthisis
Hayley Abbiss1,2,3, Garth L. Maker1,2,4, Gabrielle C. Musk1,5, Catherine Rawlinson2,4,6, Joel P.A. Gummer2,4, Patricia A. Fleming1, Jacqueline K. Phillips7, Mary C. Boyce8, John H Moncur3, Scott J Campbell3, Robert D. Trengove2,4
1School of Veterinary and Life Sciences, Murdoch University, WA, Australia, 2Separation Science and Metabolomics Laboratory, Murdoch University, WA, Australia, 3SpectralWorks, United Kingdom, 4Metabolomics Australia, Western Australian Node, WA, Australia, 5Animal Care Services, The University of Western Australia, WA, Australia, 6Faculty of Science and Engineering, Curtin University, WA, Australia, 7Department of Biomedical Science, Macquarie University, NSW, Australia, 8School of Science, Edith Cowan University, WA, Australia
First Published: Metabolomics 2016, Dublin 2016
Introduction
Nephronophthisis (NPHP) is inherited and typically presents with cysts in the kidney and liver, leading to end-stage kidney disease. Detection of NPHP and other polycystic kidney diseases (PKDs) is not sensitive or specific and management and treatment are limited to renal replacement therapy and transplantation. The Lewis Polycystic Kidney (LPK) rat has recently been classified as a model of NPHP. Using a metabolomics approach, the aim of this study was to identify early biomarkers in the blood plasma of the LPK rat. This work is complementary to a recent study which analysed kidney and liver metabolite profiles of the LPK rat.
Method
Eleven LPK and 11 Lewis age- and sex-matched control animals aged 5 to 16 weeks were used. Blood was sampled once weekly from the lateral tail vein, cell separated and plasma stored at -80°C. Metabolites were extracted with methanol and water containing 13C6-sorbitol (IS) and were dried in a rotary vacuum concentrator and then frozen. Metabolite extracts were derivatised with methoxyamine-HCl and MSTFA. For the analysis, a Shimadzu QP2010 Ultra GC-MS was used and for data analysis, AnalyzerPro®, The Unscrambler X, and SPSS were used. Compounds were matched to an in-house library of metabolites and the NIST mass spectral database.
Results
An age effect (P<0.05) was found for arachidonic acid, glutamic acid, proline, threonine and valine. A strain effect (P<0.05) was found for citric acid (Fig. 1), glycine (Fig. 1), leucine, methionine, ornithine (Fig. 1), phosphoric acid, pyroglutamic acid and serine. Age and strain effects were found for 1,5-anhydrosorbitol, cholesterol, isoleucine, linoleic acid, lysine, myo-inositol and tyrosine (Fig. 2). These findings are consistent with a previously conducted study utilising kidney and liver tissue where citric acid, serine, tyrosine and phosphoric acid were significantly different between LPK and Lewis kidney tissue and; myo-inositol, glycine, cholesterol and phosphoric acid were significantly different between LPK and Lewis liver tissue. The plasma results are generally concomitant with results from tissue however tyrosine is increased in the kidney tissue and decreased in the plasma. Of the metabolites with a significant strain or age and strain effect, citric acid, glycine and tyrosine have previously been reported as important metabolites for the PKD phenotype (1-4).
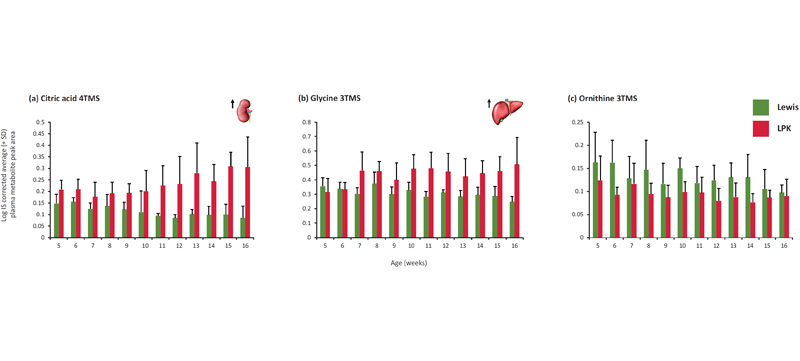
Figure 1. Comparison of Log IS corrected average (+ SD) peak area of (a) citric acid, (b) glycine, (c) ornithine in plasma between LPK and Lewis rats from 5 to 16 weeks of age. A significant strain effect was found for each of the compounds.
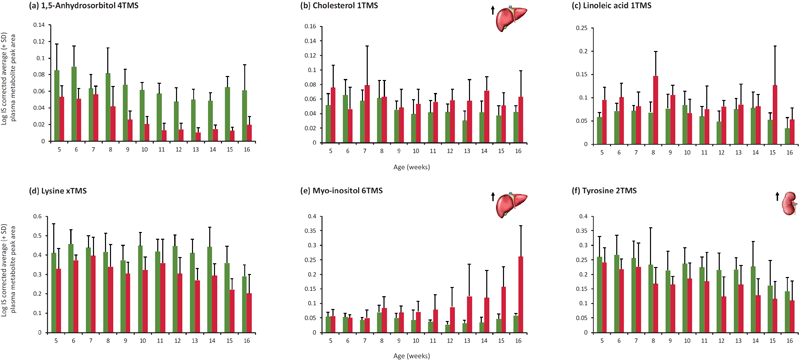
Figure 2. Comparison of Log IS corrected average (+ SD) peak area of (a) 1,5-anhydrosorbitol, (b) cholesterol, (c) linoleic acid, (d) lysine, (e) myo-inositol, (f) tyrosine in plasma between LPK and Lewis rats from 5 to 16 weeks of age. Significant age and strain effects (P<0.05) were found for each of the compounds.

Conclusion
Several LPK plasma metabolites were found to be significantly different from Lewis controls. As early markers, cholesterol, citric acid and linoleic acid may be potential candidates. Glycine and myo-inositol increased steadily as the disease progressed and 1,5-anhydrosorbitol, lysine, ornithine and tyrosine were decreased in the LPK plasma.
Acknowledgements
NCRIS Bioplatforms Australia and Shimadzu Scientific Instruments are acknowledged with thanks for supporting this work.
References
1. Gronwald W, Klein MS, Zeltner R, Schulze BD, Reinhold SW, Deutschmann M, Immervoll AK, Boger CA, Banas B, Eckardt KU, and Oefner PJ. Detection of autosomal dominant polycystic kidney disease by NMR spectroscopic fingerprinting of urine. Kidney International 79: 1244-1253, 2011. 2. Hwang VJ, Kim J, Rand A, Yang C, Sturdivant S, Hammock B, Bell PD, Guay-Woodford LM, and Weiss RH. The cpk model of recessive PKD shows glutamine dependence associated with the production of the oncometabolite 2-hydroxyglutarate. American Journal Of Physiology – Renal Physiology 309: F492-F498, 2015. 3. Taylor SL, Ganti S, Bukanov NO, Chapman A, Fiehn O, Osier M, Kim K, and Weiss RH. A metabolomics approach using juvenile cystic mice to identify urinary biomarkers and altered pathways in polycystic kidney disease. American Journal of Physiology – Renal Physiology 298: 909-922, 2010. 4. Toyohara T, Suzuki T, Akiyama Y, Yoshihara D, Takeuchi Y, Mishima E, Kikuchi K, Suzuki C, Tanemoto M, Ito S, Nagao S, Soga T, and Abe T. Metabolomic profiling of the autosomal dominant polycystic kidney disease rat model. Clinical and Experimental Nephrology 15: 676-687, 2011.